Image: Overhead view of a voltage-dependent potassium ion channel shows four red-tipped “paddles” that open and close in response to positive and negative charges. (https://www.bnl.gov/bnlweb/history/nobel/nobel_03.asp)
On the 12th of February, members of NUS Neuroscience Student Interest group convened for a seminar on Cellular Neuroscience, as part of the fortnightly seminar series hosted by group members to explore different topics in neuroscience. The seminar was hosted by Enos Goo (Year 3, Life Sciences).
The seminar started off with a presentation on the basic principles of electrophysiology and the electrochemical basis of the membrane potential that is key for the generation of Action Potentials in neurons. Enos then touched on the remarkable diversity of individual neurons in terms of the vast variations in ion channels that confer unique properties when expressed in neurons. Differential expression and distribution of such ion channels allow for unique modulation of the membrane potentials of neurons and thus contribute to their diversity.
Next, Enos shared about specific types of ion currents such as the dendritic A-type current, and the dendritic H current which confer different electrophysiological properties to neurons through the action of specific ion channels. For example, it was noted that thalamic spindles observed during sleep are a result of an interaction between a calcium ion current and an inward pacemaker dendritic H current in neuronal populations at the thalamus. This property allows for the generation of rhythmic bursts of action potentials that allows for a reduction in relaying of sensory input from sense organs to higher cortical areas for processing, since the thalamus functions as a relay centre for relaying such information (save for that of olfactory input).
It was noted that while science takes a reductionist approach in understanding the mechanisms of action in biological systems, a more global perspective yields greater insight into the more complex emergent properties of the components of such a biological system working together. An analogy was drawn to to our human society which is composed of numerous unique individuals, with not one of us being indispensable to the functioning of society as a whole. Thus, studying how a population of diverse neurons work together in neural circuits to give rise to complex emergent properties such as consciousness, the notion of morality etc., warrants significant interest.
It was also shared that back-propagated action potentials can be recorded in dendrites (albeit with a slight delay) as they too possess voltage-gated channels to support the propagation of action potentials.
The second part of the seminar involved discussing the notion of Intrinsic plasticity which involves the dynamic modulation of the electrophysiological properties of a neuron which affects their excitability and the computation of spatial and temporal summation of input the neuron receives ( as opposed to synaptic plasticity which is implicated in learning and memory) and its implications in disorders of the central nervous system. It was noted that in aging, an enhanced after-hyperpolarisation is seen at the end of action potentials and that this has been attributed to aberrant functioning of potassium ion channels which usually return the neuron to its resting membrane potential at the end of an action potential. The aberrant functioning of such channels with age could be due to the accumulation of mutations throughout life.
In epilepsy, it was noted that there was an aberrant down-regulation of potassium ion current and an enhanced persistent sodium ion current, leading to an increased intrinsic excitability of the neurons and hence the rapid burst of action potentials characteristic of an epileptic episode. In neurodegenerative diseases such as Alzheimer’s Disease (AD) and Parkinson’s Disease (PD), changes in the electrophysiological properties of the neurons have been observed prior to neuronal death (e.g. changes in ion channel expression and distribution). Moreover, degeneration of the Septal nuclei in AD leads to reduced cholinergic input to the hippocampus. This leads to an increased M current (efflux of potassium ions) and an enhanced after-hyperpolarisation of hippocampal neurons following action potentials, thus leading to the suppression of the neurons which usually fire in rapid bursts. It is noteworthy that firing in rapid bursts is an essential trait for memory coding and consolidation.
The seminar concluded with an interactive discussion on a few key topics. First, we reviewed the role of astrocytes in the ‘tripartite synapse’ and acknowledged the paradigm shift in considering the role of glial cells in the nervous system from one that views them as passively contributing to the matrix that hold neurons together in the brain to the view that they have active modulatory effects on neuronal activity (e.g. tripartite synapse’).
Next, we discussed the use of optogenetics (transfection of genes coding for light-sensitive rhodopsin activated ion channels into neurons) and considered potential applications of this technology in treating depression for example by activating serotonergic neurons instead of relying on drugs that have side-effects. However, the limitation of the technology is that light has to be directed towards the population of neurons via an optic cable (invasive). We also briefly discussed the possible physiological mechanism through which populations of neurons in migratory birds an other animals can use the earth’s magnetic field patterns to guide their migrations. We considered the possibility of using AC magnetic fields to heat (via eddy currents) a nanoparticle fused to an ion channel as a means of controlling the closure of the inactivation gate. This allows for a non-invasive way to control the activation of specific neuronal populations (a greater degree of spatial resolution).
Finally, we discussed the fact that a single nucleotide substitution on the voltage-gated sodium ion channel gene in pufferfishes allow them to be resistant to their own toxin (tetrodotoxin) which paralyses the nervous system of those who come into contact with it by inactivating the voltage-gated sodium ion channels. thus no action potential can be generated as the membrane cannot be depolarised by an influx of sodium ions. It is notable that this discovery was made by a research team based at NUS, headed by Professor Soong Tuck Wah.
While science has taught us much about the complexities of human behaviour, nothing seems to be as enigmatic as the concept of love. Love is such a powerful force that it has driven some to start great wars while inspiring others to produce marvelous works of art, poems, songs and novels. It is both rewarding and punishing, is often unpredictable and drives one to act in irrational, sometimes ridiculous ways. Einstein might be right in that this complex concept of love cannot be simply reduced to basic principles in science. However, in light of Valentine’s Day, let us review some of the insights neuroscience research has given us on how the brain handles matters of the heart.
Love, sex and other drugs
When falling in love, most people experience a rise in the levels of the stress hormone cortisol (a corticosteroid) which helps in overcoming the initial neophobia characteristic of the initial phases of starting a relationship with someone special (de Boer, et al., 2012). Many would also relate to experiencing changes in sleep patterns, a general loss of appetite and occasional mood swings when falling in love. These changes are an effect of depleting Serotonin (a neurotransmitter in the brain) levels which are inversely correlated with the levels of corticosteroids.
The brain has evolved several mechanisms to keep the passion burning after the initial period of falling in love. Most people will find it difficult to accept the idea that love is in fact an addiction. The brain is addicted to pleasure and positive affect, more specifically, it craves any activity that leads to the activation of dopaminergic pathways which trigger the brain’s reward response with the release of neurotransmitter dopamine (Insel, 2003). This pathway, known as the mesocorticolimbic pathway (see image below), originates in the ventral tegmental area (VTA) which project to the nucleus accumbens which in turn projects to the ventral pallidum and thalamus (midbrain structures as labelled below). Neurons from the thalamus then project to the prefrontal and cingulate cortex (PFC) (Everitt & Wolfe, 2002).
Research shows that addictive drugs such as cocaine are in fact addictive primarily because the activate such pathways leading to a reward response, leaving the brain craving for more (Koob & Moal, 1997). In other words, the more a particular behaviour such as taking cocaine triggers the reward centres of the brain, the more the brain seeks out such behaviour through learned operant conditioning by positive reinforcement. This then increases the occurrence of that behaviour in the future (addiction). Interestingly, in an fMRI study whereby participants were shown pictures of people they ‘liked’ versus people they ‘loved’, it was shown that the latter group elicited a greater activation of the brain’s reward pathways (Bartels & Zeki, 2000). Hence, being in love , just like drugs, triggers the same reward pathways of the brain and this leaves the brain craving for more due to positive reinforcement.
The effects of Oxytocin (OT) and arginine Vasopressin (AVP) have been extensively studied in bonding studies involving voles, comparing those that by their own nature, have monogamous relationships (prairie voles) with those that are polygamous (montane voles) (Young & Wang, 2004; Lim & Young, 2006). From such studies, it was found that blocking the release of these two hormones caused the prairie voles to be promiscuous while injection of the hormones into these voles caused them to be faithful to their partners, even when they were prevented from having sex. It would then be reasonable to expect that injecting the hormones into promiscous individuals could reduce this behaviour (many have actually suggested this as a ‘cure’ for human promiscuity!). However, injection of the hormones into montane voles which are promiscuous by nature, did not render them monogamous and it was found that the reason for this was that montane voles did not have a sufficient number of receptors for these hormones in the reward centres of their brains (Edwards & Self, 2006). Hence, they were not responsive to the reinforcing effects of oxytocin and vasopressin. This possibly suggests that certain individuals lacking the receptors for these hormones (and hence not responsive to them) are actually predisposed to exhibit promiscuity!
Oxytocin and Vasopressin are both synthesized in the brain’s hypothalamic paraventricular and supraoptic nuclei and secreted by the posterior pituitary into circulation. Both neuropeptide hormones are known to released during breastfeeding, child birth and sexual stimulation and have a neuromodulatory effect on different regions of the brain (Insel, 2010). Some of these brain regions are involved in regulating social behaviour which is then modulated as a result. Oxytocin in particular, reduces fear and anxiety related to social situations by reducing amygdala activity (Neumann, et al., 2000; Kirsch, et al., 2005), enhances social memory (Insel, 2010) and activates reward circuits in the brain involving dopamine release (Lim & Young, 2006). In fact, Oxytocin receptors have been identified in the nucleus accumbens (Insel & Shapiro, 1992) and the V1a Vasopressin receptor has been identified in the ventral pallidum (Insel, et al., 1994), both of which are part of the mecocorticolimbic pathway described earlier. Ultimately these hormones help modulate social behaviours favouring the long term maintenance of monogamous relationships through operant conditioning by rewarding such pro-social behaviours via brain’s reward system (Insel, 2010).
Quite predictably, couples with higher blood plasma oxytocin levels were found to have more positive communication behaviours (Gouin, et al., 2010), greater perceived spousal support and a higher frequency of hugs and massages (Grewen, et al., 2005). Research has also found that an individual becomes a stronger learned stimuli for oxytocin release in their partners with each successive sexual encounter with that person (Witt, 1997), proving that sex itself can be addictive. This then helps to facilitate a deeper bond between the sexual partners and increase the likelihood that they will stay together, which from an evolutionary perspective, will be essential for raising a child since the act of copulation eventually leads to begetting an offspring.
Interestingly, it seems that love can even be affected at the genetic level. High levels of a polymorphic variant of the V1a Vasopressin receptor gene, with a variation in the RS3 344 section, has been found to correlate with lower partner bonding, higher incidences of marital crises within a year and an increased likelihood of cohabitating as compared to being married to a partner in a self-report study that studied men in long-term relationships (Walum, et al., 2008).
The Social Brain
The Belongingness Hypothesis states that people have a pervasive need to form and maintain significant interpersonal relationships with others (Baumeister & Leary, 1995). In fact, research shows that the need for social interaction may be more profoundly felt than other basic needs such as hunger (Baumeister & Leary, 1995; Cacioppo, et al., 2000). Socialising with others was an essential skill for survival and reproduction in the hunter-gatherer days of human history (Baumeister & Leary, 1995). The brain has evolved over the years to become a highly social organ with specific neural networks that have been perfected over thousands of years of evolution to support this function.
One of the ways the brain has evolved to support social bond formation is by differentiating the cognitive processing of information pertaining to significant others as compared to general acquaintances. Memories and information relating to those with whom we share a significant personal relationship is processed in a unique person-by-person basis while cognitive processes for other acquaintances are stored, organised and processed on the basis of general attributional categories such as preferences, traits and duties instead of person categories (Pryor & Ostrom, 1981; Ostrom, et al., 1993). One might hypothesize that the purpose for such a distinction in cognitive processing might be to improve the speed and efficacy of the recall of information pertaining to significant people in our lives. In fact, it was reported that recall for information about both groups of people did not actually differ significantly. However, another possibility is that the specific neural networks catered to processing information about significant people may also project to emotion processing areas of the brain and thus allow for the addition of an extra layer (emotion) to the retrieval of stored information about them since relationships and attachment are closely tied to affect. This could also explain why recalling information about significant people often triggers positive emotions.
Love is blind and unconditional
The frontal cortex is the central executive centre of the brain that processes higher cognitive functions such as logical thinking, judgements, decision making and morality. In an fMRI study it was found that brain areas associated with negative emotional processing (the parietal cortex and temporal lobe) as well as other areas involved in assessing the emotions and intentions of other and other aspects of social judgements (the frontal cortex) were less active when viewing pictures of people they ‘loved’ versus pictures of their friends (Bartels & Zeki, 2000).
It then comes as no surprise that people in a relationship tend to demonstrate a self-serving bias when interpreting their partner’s outcomes in an experiment by giving them credit if they succeeded and not attributing blame to them when they fail (Fincham, et al., 1987). They also demonstrate this bias in giving their partners a more favourable interpretation of their role in causing events (causal attribution) (Craig, 1991). This probably relates to why we tend to overlook the flaws of those we are smitten over as this is probably an adaptation of the brain to aid in maintaining an existing relationship.
Researchers have also found that activity in the amygdala which is associated with fearful situations, is reduced when viewing pictures of their partners (Zeki, 2007). The suppression of judgment and increase in trust as a result of diminished fear (amygdala activity suppression) leads to increased bonding between partners and may also account for the irrational behaviour of people in love.
Rejections are painful – literally
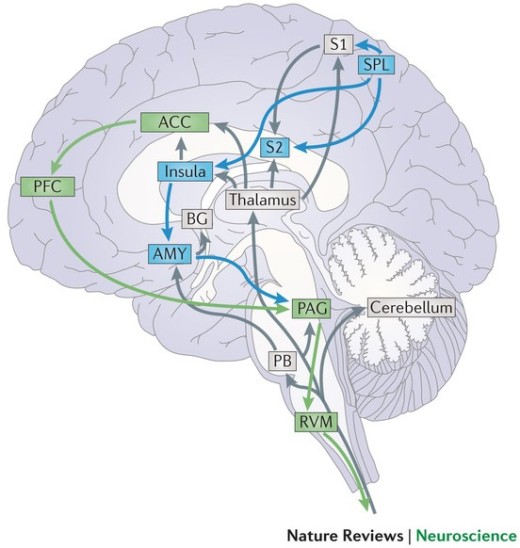
Some of us might have had the unfortunate experience of a break-up and we know that it can be an unpleasant experience. What is interesting though, is that the brain perceives the pain of social rejection the same way it would physical pain. This has been demonstrated in fMRI studies which have shown that the same brain areas, such as the anterior cingulate cortex (ACC), anterior insula (AI) and the right ventral prefrontal cortex (RVPFC), that are activated when processing the ‘affective’ or unpleasant component of physical pain, are also activated in response to social rejection (Eisenberger, et al., 2003). In fact, in one fMRI study it was found that presenting participants a picture of their ex-partners who rejected them not only activated the ACC but also triggered activity in the regions of the somatosensory cortex such as S2 which respond directly to physical sensations of pain (Kross, et al., 2011).
Image: Pain processing pathways in the brain (Bushnell et al., 2013)
It has been suggested that social system of the brain may have evolved to rely on the neural pathways for processing physical pain to indicate when social relationships are threatened, given that social connections are important for human survival (Panksepp, 1998). This further lends support to the role of operant conditioning in helping to maintain relationships. An individual learns the appropriate behaviours that will keep a relationship healthy through positive reinforcement in terms of triggering reward pathways of the brain that encourages future occurrence of such behaviour, as well as through positive punishment in terms of the pain associated with social rejection in response to behaviours that threaten relationships (Eisenberger, 2011).
The rules of attraction
An experiment involving the use of PET (positron emission tomography) to measure regional cerebral blood flow (rCBF) as a means of identifying brain areas that are more active (with more cerebral blood flow), identified that increased activity in the left insula correlated with reporting of the attractiveness of unfamiliar faces (Nakamura, et al., 1999). In what seems to be eerily similar to mind control, researchers have demonstrated that judgements of physical attractiveness can be manipulated by evoking different emotions in participants through music (May & Hamilton, 1980). It was found that evoking positive affect such that the brain’s reward pathways are activated through rock music increased perceptual judgements of attractiveness.
Anthropological research has shown that since the stone ages, people will select mates who stand out from the rest of the crowd when presented with a choice of mates of equal value (Frost, 2006). Researchers have found that the mere exposure effect, a principle in which regular exposure to a neutral of positive stimuli generally increases the liking for that stimuli (Zajonc, 1968) can be applicable to humans as well (Swap, 1977). This probably explains why some men finally win over the woman they court after some time. Either that or the women are just playing hard to get! Studies have shown that sharing a meal has a profound effect on human bonding as higher levels of oxytocin release has been measured in such settings (Wittig, et al., 2014). Hence, a nice romantic dinner on the first date might not be a bad idea after all.
In romantic situations, males use more uncommon, fancy words than they do in other situations (Rosenberg & Tunney, 2008). This could serve as a litmus test for knowledgeable women to identify if someone is trying to impress them! In one study, higher levels of fertility in women was found to be associated with lower levels of linguistic matching in their male partners in experimental setting (Coyle & Kaschak, 2012). Linguistic alignment is usually used to signal affiliation (Giles, et al., 1991) and leads to increased liking between participants in social interactions (Chartrand & Bargh, 1999; Cheng & Chartrand, 2003). However, it was noted that men paired with women in the fertile stages of their menstrual cycle, chose to use different syntactic structures in their speech and not mimic that of their partners (non-conforming behaviour) as one would expect. The men seemed to have picked up subtle subconscious cues about the female partner’s fertility and coud be presenting their non-conforming speech behaviour as a display of their fitness as a mate.
Having a sense of humour is a common feature that both men and women look for in their ideal mates, albeit with a significant difference. Men prefer women who appreciate their jokes, and not necessarily women who are funny themselves, while women prefer men who made them laugh (Bressler & Balshine, 2006). It was also observed from the study that women signal their level of attraction to their partners by the frequency of their laughter while men’s laughter was not correlated to their degree of attraction to a potential mate.
Studies have shown that men and women in a romantic relationship share fundamental differences in areas of the brain that are more active. Men, have been found to have increased activity in areas of the brain involved in integrating visual stimuli (Narumoto, et al., 2001) while women have greater activation in areas associated with memory, attention and emotion (Gray, et al., 2002; Maddock, et al., 2003; Velanova, et al., 2003). This is supported by the evolutionary view of gender priorities in looking for a mate as males look for healthy mothers to carry their child while females look for possible security and resources offered by a male mate in raising her child (Fisher, 2004).
Through better or worse
In one particular fMRI study, women who held their husband’s hands were found to have reduced activity in parts of their brain involved with processing the emotional and arousing aspects of pain when they were told to anticipate an electric shock, as compared to women who were alone (Coan, et al., 2006). Interestingly, it was found that the reduction in pain-associated activity correlated with the quality of their marriages such that happily married women had a greater degree of reduction in activity. Another observation from the study was that women who held the hands of men they did not know also showed a reduction of activity in pain processing, albeit a smaller reduction than the women who held their husband’s hands. This shows that love, or any relationship for that matter, may have a protective effect on the brain in terms of reducing the processing of unpleasant or noxious stimuli.
Conclusion
As reviewed in this article, neuroscience research may provide some insight, and at best, a fragmented view on neurological basis for some behaviours demonstrated by people in love. However, love is a complex emergent behaviour which we can never fully appreciate and artificially re-create even with the advances in knowledge.
References
Bartels, A. & Zeki, S., 2000. The Neural Basis of Romantic Love. Neuroreport, 11(17), pp. 3829-3834.
Baumeister, R. F. & Leary, M. R., 1995. The Need to Belong: Desire for Interpersonal Attachments as a Fundamental Human Motivation. Psychological Bulletin, 117(3), pp. 497-529.
Bressler, E. R. & Balshine, S., 2006. The Influence of Humor on Desirability. Evolution and Human Behavior, 27(1), pp. 29-39.
Bushnell, M.C., Ceko, M., Low, L. A., 2013. Cognitive and Emotional Control of Pain and its Distribution in Chronic Pain. Nature Reviews Neuroscience, Volume 14, pp. 502-511.
Cacioppo, J. T., Berntson, G. G., Sheridan, J. F. & McClintock, M. K., 2000. Multilevel Integrative Analyses of Human Behavior Social Neuroscience and the Complementing Nature of Social and Biological Approaches. Psychological Bulletin, 126(6), pp. 829-843.
Chartrand, T. L. & Bargh, J. A., 1999. The Chameleon Effect: The Perception-behavior link and social interaction. Journal of Personality and Social Psychology, Volume 76, pp. 893-910.
Cheng, C. M. & Chartrand, T. L., 2003. Self-monitoring Without Awareness: Using Mimicry as a Nonconscious Affiliation Strategy. Journal of Personality and Social Psychology, Volume 85, pp. 1170-1179.
Coan, J. A., Schaefer, H. S. & Davidson, R. J., 2006. Lending a Hand: Social Regulation of the Neural Response to Threat. Psychological Sciecne, 17(2), pp. 1032-1039.
Coyle, J. M. & Kaschak, M. P., 2012. Female Fertility Affects Men’s Linguistic Choices. PLoS One, 7(2).
Craig, A. A., 1991. How People Think About Causes: Examination of the Typical Phenomenal Organization of Attributions for Success and Failure. Social Cognition, 9(4), pp. 295-329.
de Boer, A., van Buel, E. M. & Ter Horst, G. J., 2012. Love Is More Than Just A Kiss: A Neurobiological Perspective On Love and Affection. Neuroscience, Volume 201, pp. 114-124.
Edwards, S. & Self, D. W., 2006. Monogamy: Dopamine Ties the Knot. Nature Neuroscience, Volume 9, pp. 7-8.
Eisenberger, N. I., 2011. Why Rejection Hurts: What Social Neuroscience Has Revealed About the Brain’s Response to Social Rejection. In: M. Brockman, ed. Future Science 19 Essays from the Cutting Edge. New York: Vintage Books, pp. 170-173.
Eisenberger, N. I., Lieberman, M. D. & Williams, K. D., 2003. Does Rejection Hurt? An fMRI Study of Social Exclusion. Science, Volume 302, pp. 290-292.
Everitt, B. J. & Wolfe, M. E., 2002. Psychomotor Stimulant Addiction: A Neural Systems Perspecive. Journal of Neuroscience, Volume 22, pp. 3312-3320.
Fincham, F. D., Beach, S. R. & Baucom, D. H., 1987. Attribution processes in distressed and nondistressed couples: IV. Self–partner attribution differences.. Journal of Personality and Social Psychology, 52(4), pp. 739-748.
Fisher, H., 2004. Why We Love: The Nature and Chemistry of Romantic Love. New York: s.n.
Frost, P., 2006. European Hair and Eye Color A case of Frequency-Dependent Sexual Selection. Evolution and Human Behavior, 27(2), pp. 85-103.
Giles, H., Coupland, N. & Coupland, J., 1991. Accomodation Theory: Communication, Context, and Consequence. In: H. Giles, N. Coupland & J. Coupland, eds. Contexts of Accommodation: Developments in Applied Psycholinguistics. Cambridge: Cambridge University Press, pp. 1-68.
Gouin, J. et al., 2010. Marital Behavior, Oxytocin, Vasopressin, and Wound Healing. Psychoneuroendocrinology, 35(7), pp. 1082-1090.
Gray, J. R., Braver, T. S. & Raichle, M. E., 2002. Integration of emotion and cognition in the lateral prefrontal cortex. Proceedings of the National Academy of Sciences of the United States of America, 99(6), pp. 4115-4120.
Grewen, K. M., Girdler, S. S., Amico, J. & Light, K. C., 2005. Effects of Partner Support on Resting Oxytocin, Cortisol, Norepinephrine, and Blood Pressure Before and After Warm Partner Contact. Psychosomatic Medicine, 67(4), pp. 531-538.
Insel, T. R., 2003. Is Social Attachment an Addictive Disorder?. Physiology and Behavior, 79(3), pp. 351-357.
Insel, T. R., 2010. The Challenge of Translation in Social Neuroscience: A Review of Oxytocin, Vasopressin, and Affiliative Behavior. Neuron, Volume 65, pp. 768-779.
Insel, T. R. & Shapiro, L. E., 1992. Oxytocin Receptor Distribution Reflects Social Organization in Monogamous and Ploygamous Voles. Proceedings of the National Academy of Sciences of the United States of Americaa, Volume 89, pp. 5981-5985.
Insel, T. R., Wang, Z. & Ferris, C. F., 1994. Patterns of Vasopressin Receptor Distribution Associated with Social Organization in Monogamous and Non-monogamous microtine rodents. Journal of Neuroscience, Volume 14, pp. 5381-5392.
Kirsch, P. et al., 2005. Oxytocin Modulates Neural Circuitry for Social Cognition and Fear in Humans. Journal of Neuroscience, Volume 25, pp. 11489-11493.
Koob, G. F. & Moal, M. L., 1997. Drug Abuse: Hedonic Homeostatic Dysregulation. Science, Volume 278, pp. 52-58.
Kross, E. et al., 2011. Social Rejection Shares Somatosensory Representations with Physical Pain. Proceedings of the National Academy of Sceinces of the United States of America, 108(15), pp. 6270-6275.
Lim, M. M. & Young, L. J., 2006. Neuropeptidergic regulation of affiliative behavior and social bonding in animals. Hormones and Behavior, 50(4), pp. 506-517.
Lim, M. M. & Young, L. J., 2006. Neuropeptidergic Regulation of Affiliative Behavior and Social Bonding in Animals. Hormones and Behavior, Volume 50, pp. 506-517.
Maddock, R. J., Garrett, A. S. & Buonocore, M. H., 2003. Posterior Cingulate Cortex Activation by Emotional Words: fMRI Evidence From a Valence Decision Task. Human Brain Mapping, 18(1), pp. 30-41.
May, J. L. & Hamilton, P. A., 1980. Effects of musically evoked affect on women’s interpersonal attraction toward and perceptual judgments of physical attractiveness of men. Motivation and Emotion, 4(3), pp. 217-228.
Nakamura, K. K. R. et al., 1999. Activation of the Right Inferior Frontal Cortex During Assessment of Facial Emotion. Journal of Neurophysiology, 82(3), pp. 1610-1614.
Narumoto, J. et al., 2001. Attention to Emotion Modulates fMRI Activity in Human Right Superior Temporal Sulcus. Brain Research. Cognitive Brain Research, 12(2), pp. 225-231.
Neumann, I. D., Torner, L. & Wigger, A., 2000. Brain Oxytocin: Differential Inhibition of Neuroendocrine Stress Responses and Anxiety-related Behaviour in Virgin, Pregnant and Lactating Rats. Neuroscience, Volume 95, pp. 567-575.
Ostrom, T. M., Carpenter, S. L., Sedikides, C. & Li, F., 1993. Differential Processing of In-Group and Out-Group Information. Journal of Personality ad Social Psychology, 64(1), pp. 21-34.
Panksepp, J., 1998. Affective Neuroscience. New York: Oxford University Press.
Pryor, J. B. & Ostrom, T. M., 1981. The cognitive organization of social information: A converging-operations approach. Journal of Personality and Social Psychology, 41(4), pp. 628-641.
Puce, A., Allison, T., Gore, J. C. & McCarthy, G., 1995. Face-Sensitive Regions in Human Extrastriate Cortex Studied by Functional MRI. Journal of Neurophysiology, 74(3), pp. 1192-1199.
Rosenberg, J. & Tunney, R. J., 2008. Human Vocabulary Use as Display. Evolutionary Psychology, Volume 6, pp. 538-549.
Swap, W. C., 1977. Interpersonal Attraction and Repeated Exposure to Rewarders and Punishers. Social Psychology Bulletin, 3(2), pp. 248-251.
Velanova, K. et al., 2003. Functional-anatomic Correlates of Sustained and Transient Processing Components Engaged During Controlled Retrieval. Journal of Neuroscience, 23(24), pp. 8460-8470.
Walum, H. et al., 2008. Genetic Variation in the Vasopressin Receptor 1a Gene (AVPR1A) Associates with Pair-bonding Behavior in Humans. Proceedings of the National Academy of Sciences of the United States of America, 105(37), pp. 14153-14156.
Witt, D. M., 1997. Regulatory Mechanisms of Oxytocin-Mediated Sociosexual Behavior. Annals of the New York Academy of Sciences, Volume 807, pp. 287-301.
Wittig, R. M. et al., 2014. Food sharing is linked to urinary oxytocin levels and bonding in related and unrelated wild chimpanzees. Proceedings of the Royal Society B Biological Sciences, 281(1778), pp. 1-10.
Young, L. J. & Wang, Z., 2004. The Neurobiology of Pair Bonding. Nature Neuroscience, Volume 7, pp. 1048-1054.
Zajonc, R. B., 1968. Attitudinal Effects of Mere Exposure. Journal of Personality and Social Psychology, 9(2), pp. 1-27.
Zeki, S., 2007. The Neurobiology of Love. FEBS Letters, 581(14), pp. 2575-2579.
On the 29th of January 2016, the NUS Neuroscience Student Interest Group conducted its first seminar titled ‘Neurons, Glia and Neurophysiology’, hosted by Sasinthiran (Year 3, Life Sciences).
Participants were provided reading material on the topic compiled from various sources two weeks prior to the seminar. At the start of the seminar, participants attempted to complete a quiz (18 questions) in groups of 3-4 members within 30 minutes.
The quiz questions were then discussed in an interactive format whereby groups had to defend and justify their answers if they differed from that of other groups.
The discussion started off with a question on the permeability of the blood-brain barrier (BBB). It was first highlighted that the components of the BBB are the tight junctions between the capillary endothelial cells, the basal lamina of the capillary and the perivascular foot processes of astrocytes (a type of glial support cell). It was noted that water and water soluble agents were able to diffuse across via the tight junctions while charged species such as sodium ions were first transported by trans-membrane protein pumps/ channels into the endothelial cells of the blood capillaries and subsequently into the brain. Lipid soluble molecules are free to diffuse through the phospholipid bi-layer of the endothelial cell membranes while larger charged molecules such as glucose ( primary energy source of the brain) and amino acids are transported by by trans-membrane transport proteins. Other solutes may also be transported by receptor-mediated endocytosis when they bind to their specific receptors on the surface of the endothelial membrane, as well as adsorptive transcytosis. It was noted that due to tight junctions between the capillary endothelial cells, leukocytes a(white blood cells) and other immune cells may not be able to cross the BBB to a great extent and hence the brain relies on a local population of cells known as the microglia (not derived from neuroectodermal lineage but from immune cell lineage) which mediate local immune function (e.g. phagocytosis and secretion of inflammatory factors such as interleukins to attract other immune cells).
Next, the discussion focused on the profile of an action potential (AP). It was noted that the refractory period (decrease in membrane potential to a value more negative than the resting membrane potential) after the peak in voltage of an AP had two parts: the absolute refractory period (lasting 1 msec due to the closing of the inactivation gate of sodium ion channels) during which no action potential can be generated followed by the relative refractory period during which a higher than usual stimulus is required to depolarize the membrane to threshold potential in order to trigger an action potential. It was also noted that the fall in membrane potential below that of resting potential because the voltage-gated potassium channels that opened at the peak of the AP were slow to close and hence allowed more potassium ions to diffuse out of the neuron, down its concentration gradient.
The discussion then looked at the factors affecting the speed of conduction along an axon and 4 factors were identified: higher temperature, wider diameter of the axon, increased myelination (insulation) and decreased length (between nodes of ranvier) all favor faster conduction. It was also noted that different classes of axons with different diameter and myelination properties exist in the body to serve their particular functions. for example, when someone hits his toes against a hard surface, the faster A-delta fibres that are myelinated conduct a sharp pain sensation while the slower unmyelinated C fibres conduct the slow, dull pain that follows.
Next, we reviewed the various gating mechanisms of ion channels: ligand-gated, mechanosensitive (e.g. stretch-activated), voltage-gated, photon-gated (responds to light) or even ungated. We briefly discussed the recent development of optogenetics in which genes coding for a light-sensitive receptor has been transferred and expressed in select populations of mice neurons to modulate their behavior when that region is exposed to light via an optic fiber cable.
Our discussion then moved on to define the terms that describe a cell membrane’s potential. Depolarisation represents a rise in membrane potential from a negative resting potential towards the potential of 0mV (same internal potential as outside of the cell). Repolarisation represents a drop in membrane potential from 0mV to a more negative potential. Hyperpolarisation refers to a drop in membrane potential to a value more negative than that of the resting potential.
Next, we had a brief review of the activity at the neuromusccular junction (NMJ) that results in the contraction of skeletal muscle fibres for movement. It was noted taht acetylcholine was the primary neurotransmitter released at the NMJ.
The discussion then moved on to briefly touch on the fact that groups of axonal tracts that run up and down the spinal cord are arranged in columns. It was noted that such tracts are called projections when observed in the brain.
We then proceeded to discuss the factors responsible for the establishment and maintenance of the action potential: the unequal distribution of ions inside and outside the neurons (sodium, chloride and calcium ions are concentrated outside the cell while the inside of the cell has a higher concentration of potassium ions and other large non-difussible anions that also contribute to the negative resting membrane potential), the action of the sodium-potassium ATPase pump which pumps out 3 sodium ions and pumps in 2 potassium ions against their concentration gradient (thereby resulting in the net loss of one positive ion) as well as the free diffusion (leakage) of potassium ions out of the cell through leaky potassium channels.
Next, the discussion focused on the role of glia in the nervous system. It was noted that there has been a paradigm shift from considering glia simply as cells that form the matrix that hold the neuron together to one that accepts their active role in the nervous system. For example, astrocytes have been found to form a ‘tripartite synapse’ with post- and pre-synaptic neurons and even modulate and synchronize their communication through the release of gliotransmitters (visualised trhough the observation of signature calcium waves in the astrocytes which lead to the fusion of vesicles containing gliotransmitters to the astrocyte membrane to release the gliotransmitters). Microglia have the potential to differentiate into astrocytes or neurons, modulate local immune response through phagocytosis and release of cytokines such as interleukins to attract other immune cells (inflammation) as well as guide young neurons during neuronal migration in early development. Schwann cells (in the PNS) and oligodendrocytes (in the CNS) form myelin sheaths around axons to nourish the cells and at the same time speed up the conduction of action potentials. It was noted that the key difference between neurons and glia is that glia do not generate action potentials. It was noted, however, that a population of the pacemaker cells in the sinoatrial (SA) and atrioventricular (AV) nodes of the heart are the only other cells outside of the nervous system that are capable of generating their own action potentials.
The next question revisited the profile of the action potential and it was noted that the rapid depolarisation of the membrane was due to the rapid influx of sodium ions through their voltage-gated channels (more open upon membrane potential reaching threshold potential), driven by both their electrical and chemical gradients into the neuron.
Next, we looked at the summation of input from pre-synaptic neurons in a post-synaptic neuron that could either lead to to an EPSP (excitatory post-synaptic potential) or an IPSP (inhibitory post-synaptic potential). It was noted that if the excitatory effect is greater than the inhibitory effect but less than the threshold of stimulation, the result is a subthreshold EPSP. Furtehrmore, if the excitatory effect is greater than the inhibitory effect and reaches or surpasses the threshold level of stimulation, the result is a threshold or suprathreshold EPSP and one or more nerve impulses. Alternatively, if the inhibitory effect is greater than the excitatory effect, the membrane hyperpolarizes, resulting in inhibition of the postsynaptic neuron and the inability of the neuron to generate a nerve impulse.
Subsequently, we went on to review the fact that stimuli intensity is encoded in the frequency of action potentials which are of equal magnitude (amplitude). The higher the stimuli intensity, the higher the frequency of the action potential.
We also identified that the falling of the membrane potential towards resting membrane potential was due to the opening of voltage-gated potassium channels at the peak of the action potential, leading to an efflux of potassium ions out of the cell, down its concentration gradient. As noted before, the slow closure of these channels results in the drop in membrane potential past that of the resting membrane potential (refractory period).
Next, we briefly discussed the experiment conducted by German physiologist Otto Loewi which proved that neurons communicated through chemical messengers that we now know to be neurotransmitters. He stimulated one frog’s heart, collected fluid around it, transferred it to another frog’s heart, and saw change in its heart rate.
We then moved on to briefly discuss a thought experiment and concluded that if we wanted to cause the pre-synaptic terminal of an axon to release its neurotransmitter without an action potential, we could inject calcium into its pre-synaptic terminal. This is because of the fact that the influx of calcium into the pre-synaptic terminal (usually when an action potential reaches the pre-synaptic terminal and depolarise the membrane, causing the opening of voltage-gated calcium ion channels) results in teh fusion of synaptic vesicles containing neurotransmitters to the pre-synaptic membrane.
Next, we discussed the differences between metabotropic and ionotropic receptors. Ionotropic receptors have a pore which opens/ closes to directly control the passage of ions while metabotropic receptors when activated, trigger an intra-cellular signalling cascade to cause an ion channel to open/ close in response (indirect control of movement of ions). Thus ionotropic receptors have fast acting effects and and its action is short-lived while metabotropic receptors have slower effects that last longer (due to signal amplification in the signalling cascade).
The seminar concluded with the discussion of the learned concepts in more challenging application questions. Firstly, we discussed the scenario whereby an action potentaial is triggered at same time from both directions of an axon (in both the orthodromic and antidromic directions). In this case, we concluded that when both action potentials meet at a point along the axon, they will not propagate any further. This is because the region just behind the each action potential is undergoing a 1 msec absolute refractory period in which no action potential can be generated since the voltage -gated sodium ion channels have their inactivation gates closed for that period.. Thus both the orthodromic and antidormic action potentials cannot proceed ahead in their direction of propagation.
There was also a short sharing on the role of GABA as an excitatory neurotransmitter in the neurons of fetuses (due to the high intracellular chloride ion concentration). The role of Glutamate as both an excitatory and inhibitory neurotransmitter in the retina was also discussed and it was noted that the effect of neurotransmitters is dependent on the type of receptor it binds to (metabotropic or ionotropic).
With over 90 years of experience, the School of Psychology and Clinical Language Sciences at the University of Reading, houses a range of sector leading programmes. We offer MSc and PhD programmes, as well as international scholarship opportunities for students. The school is unique in housing dementia, anxiety and depression research clinics, and is also home to a purpose built neuroimaging facility (EEG, TMS and fMRI).
The University of Reading is a leading research intensive institution in the UK, situated on a beautiful green campus just half an hour away from London Heathrow Airport, and close to all major transport links. Please don’t hesitate to contact us, or visit the website for further information http://www.reading.ac.uk/psychology/.
The Bernstein Center for Computational Neuroscience Berlin and the TU
Berlin invite applications for *6 fellowships* of the Research Training
Group “Sensory Computation in Neural Systems” (GRK 1589/2, https://www.eecs.tu-berlin.de/grk_15891/).
The *scientific program* of the research training group combines
techniques and concepts from machine learning, computational
neuroscience, and systems neurobiology in order to specifically address
sensory computation. Doctoral candidates will work on interdisciplinary
projects investigating the mechanisms of neural computation, address the
processes underlying perception on different scales and different levels
of abstraction, and develop new theories of computation hand in hand
with well-controlled experiments in order to put functional hypotheses
to the test.
The training group offers structured supervision complemented by a
teaching and training program. Each student will be supervised by two
investigators with complementary expertise and will be associated with
the Bernstein Center for Computational Neuroscience Berlin
(https://www.bccn-berlin.de/) a leading research center dedicated to the
theoretical study of neural processing.
Candidates are expected to hold a Masters degree (or equivalent) in a
relevant subject (e.g., neuroscience, cognitive science, computer
science, physics, mathematics, etc.) and have the required advanced
mathematical background.
Candidates selected in the first application step will be invited for
lab visits and an interview, expected to take place in June 2015. The
*fellowships of 1468 €/month* – with additional children allowances if
applicable—will be granted for up to three years.
Master’s Program
The tuition-free Master program in Computational Neuroscience offers *15
places* per year, has a duration of 2 years and is fully taught in English.
The *curriculum* is subdivided into ten modules, whose content includes
theoretical neuroscience, programming, machine learning, cognitive
neuroscience, acquisition, modelling, and computational analysis of
neural data, with a strong focus on a complementary theoretical and
experimental training. Three lab rotations and a Master’s thesis are
accomplished in the second year. The aim of the program is to provide
the students with an interdisciplinary education and an early contact to
the neurocomputational research environment.
*Requirements* BSc or equivalent degree in a relevant subject (typically
in the natural sciences, in an engineering discipline, in cognitive
science, or in mathematics), certificate of English proficiency, proof
of sufficient mathematical knowledge (at least 24 ECTS credit points).
The NUS Neuroscience Student Interest Group is an academic student group which aims to bring together like-minded undergraduates across the faculties at the National University of Singapore (NUS), with a passion for neuroscience, in a collaborative learning environment.
The interest group hosted its Welcome Tea on 4th January to inaugurate the interest group and to orientate members to the upcoming schedule of activities. The slides from the Welcome Tea are available here: Welcome Tea
Our activities include bi-weekly Journal Club sessions during which we will engage in an interdisciplinary discussion on current research in the field, attending symposiums and seminars organized by the various departments at NUS and other research institutions, learning about the research projects of peers who are undertaking their FYP or UROPS in the field, and possibly even organizing events and activities for the university community and beyond.
This interest group will be a great opportunity for undergraduates to get an early exposure into the field and meet like-minded peers who could be potential collaborators, especially if they are considering a post-graduate career/ education in neuroscience. We hope to cultivate a community of collaborative learning to supplement our undergraduate education with exposure to the field, especially since neuroscience is not offered as an area of specialization/ major at NUS.
The possible topics that we will be covering for the upcoming bi-weekly Journal Club meetings can be found here: Journal Club Topics
The list of topics are based on chapters from a free, peer-reviewed textbook (Neuroscience in the 21st Century: From Basic to Clinical), which will serve as the primary resource for the Journal Club. Members will be split into groups of 3-5 based on their interest in the topics available.
In brief, groups will have to prepare a review presentation of the topic selected not exceeding 30 minutes using material found in the textbook as well as other relevant primary literature. The rest of the members in the interest group are encouraged to read up on the relevant topics/ chapters and come up with their own ideas either from their own experience or literature review. After the presentation, the presenting group will facilitate an open-floor discussion in the remaining time (they may choose to structure it in any way they prefer) and invite opinions and ideas from the other members who will share relevant ideas and insights from their own disciplines. In this way, we hope that each session provides a multi-disciplinary view of the topics discussed, to all in attendance.
As part of our ad-hoc initiatives, this blog was set up with the aim of serving as a platform for the interest group to share about our activities, archive the knowledge that is shared during our discussions as well as to encourage science-related writing among our members. Our team of writers will periodically publish articles featuring events of the interest group, reviews of recent neuroscience research articles of interest as well as feature interviews with neuroscience researchers in Singapore.
Do follow our blog to keep updated on our events and to join us in this exploratory journey in discovering more about the brain!
Sasinthiran
President, NUS Neuroscience Student Interest Group




![_20160209_181639 [2344380].JPG](https://nusneurointerest.files.wordpress.com/2016/02/20160209_181639-2344380.jpg?w=441&h=576)


{kind=link}