
Written by Yingchen
Edited by Xin Chen, Sasinthiran and Keshiniy

On the 22nd of February 2016, members of NUS Neuroscience Student Interest group attended a workshop on Neuroimaging hosted by A*STAR-NUS Clinical Imaging Research Centre (CIRC).
The Workshop started with an introductory lecture by Dr John Totman, Head of Imaging Operations at CIRC. He recounted the history behind various imaging techniques such as Ultrasound, X-Ray, CT, PET and MRI, and gave a brief overview on the science behind these techniques, their limitations and common uses, for example, in the field of nuclear medicine.
One of the most common imaging techniques, ultrasound makes use of the Doppler Effect. Ultrasound waves are emitted from the probe, which then detects the reflection of those waves off anatomical structures to construct an image of the structure; similar to echolocation used by dolphins and bats. Ultrasound is not commonly used in neuroimaging because sound waves cannot penetrate the skull.
Another commonly used technique, X-ray works on the principle that structures in the body attenuate x-rays that are projected on one side of the body such that the rays emerging from the other side expose a sheet of film to produce an image reflecting the body structures. However, the images produced are two-dimensional and thus may miss out certain structural details. As a result, multiple images at different sections of the body are required for a more accurate representation of the body structures.
A variant of X-ray imaging is Computerized Tomography (CT). It uses a rotating x-ray emitter and detector to provide a 3D reconstruction of anatomical structures. It is commonly used in neuroimaging to provide the structural information that aids in medical practices, such as detecting tumours and aneurysms. The grey and white matter of the brain can be distinguished clearly on CT images.
Another technique, Positron Emission Tomography (PET) requires an ingestible tracer consisting of glucose in conjugation with radioactive fluorine, which is prepared in the expensive cyclotron reactor by accelerating and colliding sub-atomic particles. Other positron-emitting radionuclides such as oxygen might also be used depending on how long the effect is required to last. When the radioactive substance decays, positrons are released which collide with the electrons released by the PET scanner, producing two gamma rays which scatter in exactly opposite directions. The PET scanner detects these two gamma rays and computes the average distance at which the rays originated to indicate the location of the anatomical structure. PET can be used for functional as well as anatomical imaging. The principle behind anatomical imaging is that cancer cells take up glucose rapidly, but fludeoxyglucose (FDG) cannot be easily metabolised and thus accumulate in these cells and can be detected.
SPECT, which stands for Single-photon Emission Computerized Tomography, is a combination of PET and CT. PET provides metabolic information while CT provides structural anatomical landmarks such as bones to give relevance to the metabolic activity reported by PET scans.
After Dr Totman’s lecture, Ms Caroline Wong, a Research Officer at CIRC, introduced us to Functional Magnetic Resonance Imaging (fMRI), which is the most commonly employed imaging tool for functional studies in neuroscience research. fMRI provides both structural and functional information of the brain. The basic unit of fMRI images is the voxel, which is a 3D pixel with modifiable size and length. Smaller the voxel, greater is the time required to acquire it, but more detailed the image would be, which is analogous to having thinner slices: many voxels make up a slice, and many slices make up the volume.
The science behind MRI is that protons have different spins in random directions, which align when placed in a magnetic field. When the MRI scanner emits radio frequency pulses, the protons are tilted out of alignment from each other. When the radio frequency pulses stop, the protons lose energy and return to their baseline aligned state, thereby releasing electromagnetic waves that are detected by the scanner.
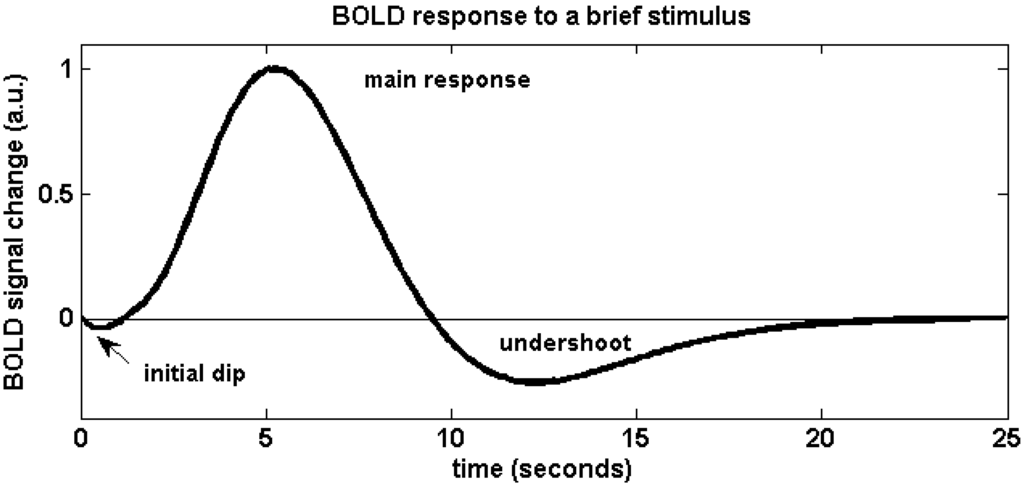
In fMRI practice, active areas of the brain receive more oxygenated blood flow due to the dilation of local cerebral blood vessels. The protons in oxygenated haemoglobin and deoxygenated haemoglobin have different rates at which they return to their baseline aligned state. Moreover, oxygenated blood is diamagnetic, does not distort the surrounding magnetic field and thus there is no signal loss. In contrast, deoxygenated blood is paramagnetic, distorts the surrounding magnetic field and thus there is signal loss. These differences are used to highlight brain areas that are active during a task, since active areas are marked by a higher level of oxygenated blood flow. A more detailed relation is displayed by Figure 2, the hemodynamic response function profile.

In Figure 2, the initial dip is due to blood oxygen taken in by active brain areas from surrounding blood vessels. The rapid rise is due to overcompensation of blood flow due to dilation of blood vessels, which usually lasts for around 4-8 secs, but the exact time period depends on the brain area. The post-stimulus undershoot is due to elastic recoil of expanded blood vessels. Meanwhile, voxel colour changes with the time course of the curve.
Typical fMRI task designs include the block design, which consists of periods of rest between periods of activity; the slow event-related (ER) design, which has been phased out as it is inefficient; the rapid counterbalanced ER design, which is the fastest; and the mixed design, which consists of both block and ER. One difficulty with fMRI is that it requires minimal movement of non-task related areas of the body since movements may create “noises” that confound the results. Another difficulty is that brain functions are not localised to particular areas; meanwhile, the same area may be involved in many different functions. Notably, synchronisation between multiple brain areas that process the same cognitive properties have been seen to be out of synchrony in patients with neuropsychological disorders such as schizophrenia and bipolar disorder. Thus, careful interpretation of neuroimaging results is necessary.
After the presentations, we were invited to visit the fMRI laboratory, and witnessed several interesting phenomena such as the strong force of attraction between metal objects and the fMRI scanner, and the fact that conductors would fall due to gravity when suspended, albeit at a slower than expected rate in the fMRI scanner chamber due to electromagnetic induction. Towards the end of the workshop, Ms Caroline Wong also indicated that Dr Qiu Anqi from NUS Computational Functional Anatomy Lab was looking for research assistants. Details can be found in this link: http://www.bioeng.nus.edu.sg/cfa/.
Within the two-hour session, the workshop provided us with an essential foundation for appreciating the various neuroimaging techniques used in neuroscience research and clinical practice. We would like to thank the Clinical Imaging Research Centre (CIRC) for accommodating our group and conducting this workshop.
Reference:
“MDPI Open Access Publishing”, MDPI AG, 1996-2016. Accessed 7th March 2016. (http://www.mdpi.com/materials/materials-04-01941/article_deploy/html/images/materials-04-01941-g001-1024.png.)

![_20160209_181639 [2344380].JPG](https://nusneurointerest.files.wordpress.com/2016/02/20160209_181639-2344380.jpg?w=441&h=576)


{kind=link}